
Les Transmembranaires
Les Transmembranaires sont des protéines incorporées et traversant la membrane cellulaire qui est une barrière sélective entourant la cellule, composée principalement d’une double couche, appelée bicouche phospholipidique, dans laquelle sont intégrés les protéines transmembranaires, associées étroitement via leur région hydrophobe centrale, exposant leurs extrémités C-terminale hydrophiles de part et d’autre de la bicouche au contact des milieux aqueux, et l’autre extrémité N-terminal hydrophobe tournée vers les lipides.
Le Transport
Le Transport transmembranaire est le mouvement de substances à travers la membrane plasmique par le biais de protéines qui agissent comme des canaux ou des transporteurs. Le Transport vésiculaire est un autre processus de déplacement de substances à l’intérieur des poches sphériques appelées vésicules, fusionnant avec la première couche puis la deuxième couche tout dépend du sens du déplacement dans la membrane. Toutes les cellules doivent échanger des substances avec leurs environnement pour maintenir leur homéostasie. Ces échanges se font principalement via des transports transmembranaires et vésiculaires. Ces mécanismes permettent le passage de molécules nécessaires comme les nutriments et l’expulsion des produits par le métabolisme intracellulaire.
Les Types de transporteurs
Les transporteurs sont des pores situés dans la bicouche lipidique. Ils sont formés par de grands complexes protéiques intrinsèques traversant la membrane, reliant le cytosol à la matrice extracellulaire. Leurs propriétés principales est la sélectivité et le contrôle de façon très fine et le transport des métabolites.
Canaux : Les Canaux sont des protéines-canal assurant le transport passif de molécules à travers la bicouche. Le passage à travers un canal suit les lois de la diffusion, cependant, parfois elles peuvent être plus ou moins sélectives. Les canaux peuvent aussi se fermer et s’ouvrir en fonction de différents stimulus, qu’il soit électrique, chimique ou mécanique. Le potentiel de la membrane est soumis à la sensibilité de certaines cellules et certains signaux extérieurs.
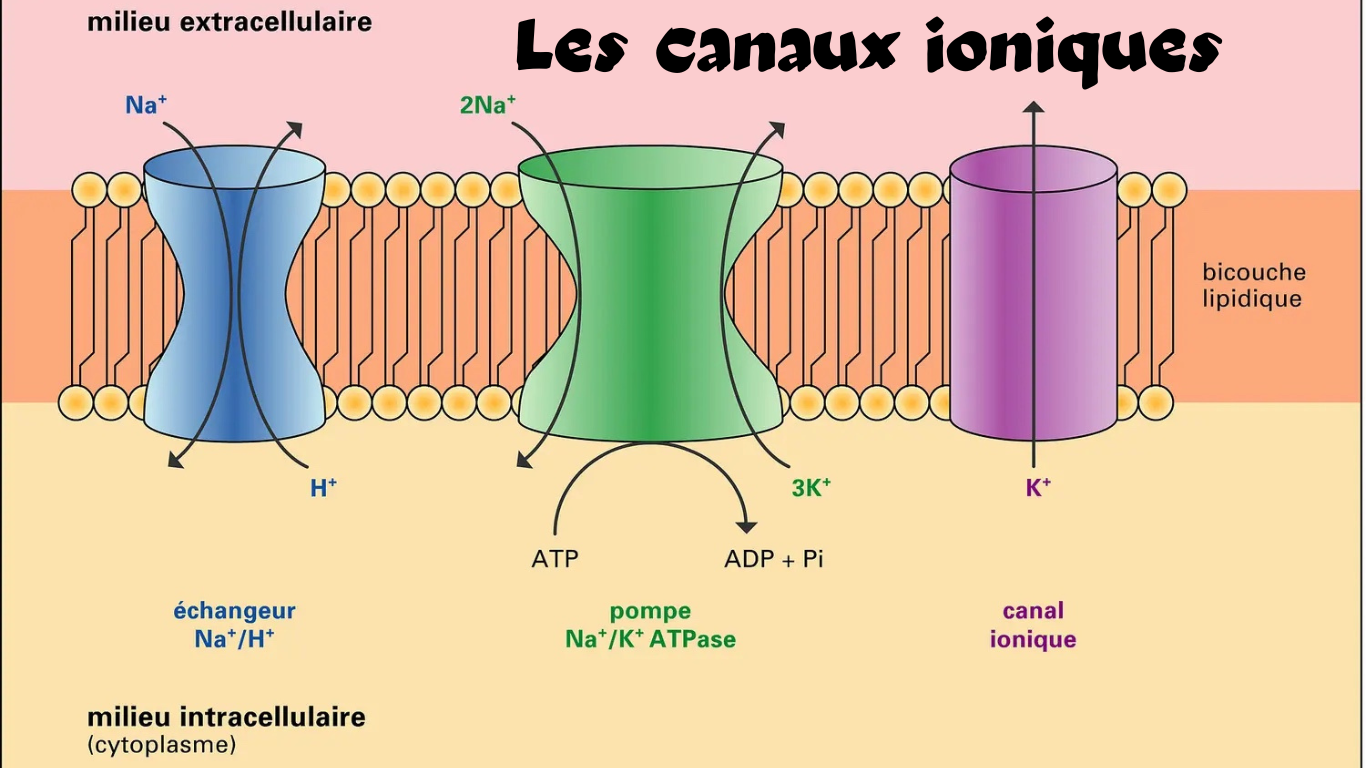
Les canaux ioniques: Les canaux ioniques principaux sont les canaux sodiques (Na+), potassiques (K+), chloriques (Cl-) et calciques (Ca++). Certains canaux sont spécifiques pour un ion donné, mais plusieurs canaux permettent le passage de plus d’une sorte d’ion.
Les pompes: Les pompes sont des canaux mais avec la particularité de fonctionner à l’inverse des canaux ioniques, les pompes Na-K (sodium-potassium) travaillent à contre-courant en retournant de façon continue les ions, là où ils sont en grande concentration afin de maintenir les dosages inégales d’ions de part et d’autre de la membrane et à la repolarisation. Les pompes Na-K sont en fait des canaux ioniques particuliers fonctionnant à très haute consommation d’énergie.
Transport passif
En biologie, le transport passif désigne le passage d’un ion ou d’une molécule à travers la membrane sans apport d’énergie, contrairement au transport actif, lui consomme de l’énergie. Dans le transport passif, les porines sont des pores constitués d’une ou plusieurs sous-unités protéiques transmembranaires. Le passage à travers la membrane nécessite peu de changements de configuration de la protéine de transport.
Les transporteurs actifs
Les transporteurs actifs sont très importants dans la cellule, car c’est eux qui concentrent le calcium du cytoplasme vers le réticulum endoplasmique, maintient ainsi la concentration cytosolique libre. le calcium est utilisé par de nombreux récepteurs comme signal pour prévenir la cellule de la présence de sa molécule activatrice sur son site actif. Les transports actifs ne fonctionnent qu’en présence d’énergie métabolique en présence de l’ATP, sans cette source d’énergie rien n’est possible contre les forces du gradient électrochimique.
Les co-transporteurs et les pompes sont formés de protéines sans pore apparent. Le passage d’ions ou de petites molécules nécessite donc un profond changement de configuration. Ce transport est beaucoup plus lent mais permet aux solutés de traverser contre leur propre gradient électrochimique. Les co-transporteurs et les pompes transfèrent les molécules de soluté à travers la membrane selon le principe d’une réaction enzyme-substrat, tel que, E+S <–> ES –> E+P; (E étant le transporteur, S le soluté dans le compartiment initial, et P le produit de soluté transféré dans le compartiment final). Cette approche permet la détermination de constantes physicochimiques Vmax et Km (Vmax pour vitesses max et Km pour constance de Michaëlis Menten) caractéristiques de chaque transporteur. Les transporteurs possèdent un ou plusieurs sites de fixation spécifiques du substrat. Cependant, à la différence d’une réaction enzyme-substrat, le soluté transporté n’est pas modifié. E+S→k1←k–1ES→k2 E+P.
![]() Les Transporteurs conduisant les molécules contre leur gradient électrochimique, ne nécessite pas forcément l’hydrolyse de l’ATP. Il existe de nombreux cas où l’énergie est fournie par un ion ou une autre molécule qui suit son gradient électrochimique. Ce phénomène s’appelle transport couplé ou Co-transport, car il couple un canal ionique à une pompe membranaire et utilise l’énergie de l’un pour activer l’autre. Selon le sens de déplacement respectif de deux molécules on parle de Symport, c’est à dire, l’ion et la molécule transportés traversent la membrane dans le même sens. Quand les deux espèces chimiques se déplacent en sens inverse, là il s’agit du transport Antiport. Ces transports couplés sont très utilisés par la cellule pour récupérer les molécules nécessaires à son métabolisme dans le milieu extérieur.
Les Transporteurs conduisant les molécules contre leur gradient électrochimique, ne nécessite pas forcément l’hydrolyse de l’ATP. Il existe de nombreux cas où l’énergie est fournie par un ion ou une autre molécule qui suit son gradient électrochimique. Ce phénomène s’appelle transport couplé ou Co-transport, car il couple un canal ionique à une pompe membranaire et utilise l’énergie de l’un pour activer l’autre. Selon le sens de déplacement respectif de deux molécules on parle de Symport, c’est à dire, l’ion et la molécule transportés traversent la membrane dans le même sens. Quand les deux espèces chimiques se déplacent en sens inverse, là il s’agit du transport Antiport. Ces transports couplés sont très utilisés par la cellule pour récupérer les molécules nécessaires à son métabolisme dans le milieu extérieur.
Protéines de transport
Les transporteurs sont des protéines transmembranaires qui comportent un site de fixation à l’ion ou/et à la molécule à transporter, ils subissent alors un changement de conformation pour déverser l’ion ou la molécule fixé de l’autre côté de la membrane.
Le Passage par diffusion simple: est un mode de transport ne fait pas intervenir de protéines membranaires. Il est limité aux gaz tel que, le N2, l’O2, le CO2, et le NO, et les molécules lipophiles comme les hormones stéroïdiennes et thyroïdiennes, l’urée, l’éthanol et dans certaines limites du H2O.
Le Passage par protéines de transport: La diffusion par un transporteur augmente très largement la vitesse et la sélectivité de transport par rapport à la diffusion simple. Le transporteur de glucose, comme la perméase GLUT-1 illustre bien ces deux aspects. La diffusion par un transporteur permet également de transporter les solutés contre leur gradient chimique et électrique consommant de l’énergie, on parle alors de transport actif. Le transport actif permet le maintien de concentrations de solutés différentes de part et d’autre de la membrane.
Comportement des molécules chargées
La sélectivité de transport dépend de la nature des canaux. Pour les canaux ioniques le passage semble être déterminé par deux paramètres, les forces électrostatiques à l’intérieur du canal et le diamètre du port. L’intérieur des canaux anioniques de chlorure CI-, est faiblement positif et celui des canaux cationiques potassique, K+ est faiblement négatif. L’ion est alors attiré par la faible charge opposée et se positionne au milieu du canal dans un environnement électrostatique favorable.
Pour la majorité de canaux ioniques, les ions qui entrent perdent la plupart des interactions électrostatiques avec l’eau, ils établissent de nouvelles interactions avec le pore. Ils traverseront complètement à la suite de l’entrée d’autres ions de même charge qui le repousseront vers l’autre côté du canal. Le mécanisme proposé justifie le principe du passage des ions en fonction de leur gradient de concentration. La différence de potentiel électrique au travers la membrane joue aussi un rôle important, parce qu’elle peut s’opposer à la sortie des ions.
Maintien des concentrations ioniques
L’ATPase Na+/K+ ou pompe NA+/K+, La concentration en potassium-K+ est typiquement 30 à 40 fois plus élevée à l’intérieur des cellules qu’à l’extérieur, alors que la situation est inversée pour le sodium-Na+. Ces différences sont engendrées et maintenues par une ATPase de la membrane plasmique, qui se comporte comme une pompe expulsant activement 3 ions Na+ vers l’extérieur de la cellule et important deux ions K+ vers l’intérieur.
La pompe ATPase Na+/K+: comme la H+/K+_ ATPase et la Ca+/K+ _ ATPase, est une enzyme pompe, permettant le transfert transmembranaire des cations sodium-Na+ et potassium-K+. Comme la plupart des autres pompes, elle est constituée de trois protéines étroitement liées (sous-unités α,β,λ, respectivement d’un poids moléculaire de 110 kDa, 40 kDa et 8 kDa). Son mécanisme moléculaire de transport exige trois étapes, la fixation des ions à haute affinité, transfert des ions au travers de la membrane, et réduction d’affinité qui permet la libération des ions. Le gradient Na+/K+ généré de part et d’autre de la membrane est essentiel au fonctionnement de la cellule. Il est impliqué dans diverses fonctions, par, régulation du pH, régulation du volume cellulaire, transport de nutriments tels que glucose et certains acides aminés. Le gradient Na+/ K+ est important pour la transmission du signal dans le système nerveux par potentiel d’action.
La différence de potentiel
La différence de potentiel transmembranaire, ou potentiel de membrane, d’une cellule animale est proche de -70 milli Volt, la face cytoplasmique étant chargée négativement par rapport à la face externe. Le potentiel de membrane est le résultat de mouvements ioniques transmembranaires. Ces mouvements sont la conséquence d’une distribution inégale de part et d’autre de la membrane des ions et les macromolécules chargées, tel que, les glucides complexes, les nucléotides et les protéines. Cette distribution est elle-même la conséquence de transports transmembranaires actifs avec une contribution majeure de l’ATPase Na+/K+.
Dans un état repos c’est le mouvement de K+ au travers de la membrane qui prédomine, parce qu’il y a plus de canaux potassiques que de canaux sodiques ouverts. En conséquence, la valeur du potentiel de repos est essentiellement déterminée par le mouvement de K+. Grâce à sa concentration intracellulaire très élevée, le K+ sort de la cellule en polarisant la face cytoplasmique négativement par rapport à la face externe. Le potentiel ainsi créé s’oppose au mouvement suivant de K+ au travers de la membrane, c’est-à-dire que le gradient électrochimique de K+ diminue. Sans la présence de Na+, le potentiel atteint la valeur de moins de 90 milli Volt, une valeur pour laquelle il y a équilibre entre les deux forces. Cependant, le potentiel de membrane créé par le K+, induit une augmentation considérable du gradient électrochimique du Na+, ce qui provoque un flux entrant de Na+ de plus en plus important. A un moment donné, il s’installe un équilibre dynamique où il y autant de K+ qui sortent que de Na+ qui entrent, c’est le potentiel de membrane de repos.
Sources & références


